Chapter 10 - Activation and Function of T and B cells
1. CD 4 (+) T-cells become activated by antigen presenting cells (APC's). Naive CD4(+) cells are activated by dendritic cells. Memory CD4(+) cells interact well with macrophages.
Figure 10.1 shows the basic model of how dendritic cells interact with CD4(+) T-cells:

Dendritic cells are phagocytic cells that also have sets of pattern recognition receptors called Toll-like receptors (TLRs).
The engagement of TLRs by microbial products causes:
· The release of a number of cytokines and chemokines, many of which are pro-inflammatory
· The maturation of the dendritic cells into efficient antigen presenting cells
o Increase MHC class II molecule expression
o Increase B7 expression
o Migration of the dendritic cell to T-cell areas of the lymph nodes
Figure 10.2 is a diagram of the Immunological Synapse
This indicates all of the receptor combinations that must come together in order to acivate CD4(+)T-cells
The ultimate result of the interaction of all of these receptor combinations is the activation of intracellular signaling cascades as indicated in Fig. 10.3

· Tyrosine kinases (src family = fyn and lyk) phosphorylate “immunoreceptor tyrosine-based activation motifs” or ITAMS on the intracellular parts (cytoplasmic tails) of the various receptors.
· New tyrosine kinases (syk family = ZAP-70), attracted to the newly phosphorylated cytoplasmic tails, get phosphorylated and activated.
· ZAP-70 then phosphorylates and activates adaptor molecules which attract, bind and activate a large group of enzymes (such as phospholipase C-g and protein kinase C)
· This in turn causes the activation of a group of cytoplasmic transcription factors (NF-kB, NF-AT and AP-1). These transcription factors move into the nucleus of the newly activated T-cell and activate the transcription of “immune response genes”, causing the growth and proliferation of the T-cell as well as the synthesis of cytokines
(Note that certain substances such as superantigens and lectins (such as concanavalin A, pokeweed mitogen and phytohemagglutinin) can short circuit the above mechanism through promiscuous binding. Such substance are often referred to as polyclonal activators or mitogens.)
Figure 10.5 summarizes current understanding about the differentiation of CD4(+) T-cells after activation. Note that these cells can develop down different paths which are identified by the constellation of cytokines which they produce. Note also that the differentiation pathway is influenced (determined) by the cytokines present at the time of initial antigen activation:
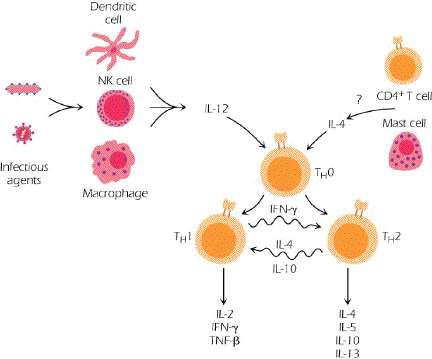
· Thus if there is plenty of IL-12 around at the time of activation, then TH1 cells develop. On the other hand, if there is plenty of IL-4 around, then TH2 cells develop.
o TH1 cells are characterized by the production of IL-2, IFN-g and TNF-b (see pg 153).
· These cells help activate “cell-mediated immunity” – primarily CD8(+) T-cells, NK cells and macrophages. IgG antibody responses are augmented by TH1 cells too.
o TH2 cells are characterized by the production of IL-4, IL-5, IL-10 and IL-13 (see pg 153)
· These cells are important for efficient antibody production, particularly for the production of IgE
o TH1 and TH2 cells can inhibit each other.
T and B cells cooperate in the evolution of the immune response to an antigen through the process of linked recognition
Note, as shown in fig 10.6, that B-cells have the ability to capture and present antigen. Because this happens through the B-cell receptor, this is “specific” capture and presentation.

This allows the efficient presentation of antigen to CD4(+) T cells by B-cells that already have the ability to make antibody to that antigen.
Figure 10.7 shows the various receptors that engage between B-cells and T-cells. Note that there are many parallels to what we have seen already with the dendritic cell model.
Isotype switching will require both the engagement of CD154 with CD40 and the production of cytokines by the T-cell.
Cytotoxic T-cells – CD8(+) T-cells
Figure 10.8 diagrams 2 things: a) How CD8(+) T-cells get activated and b) How they kill target cells.
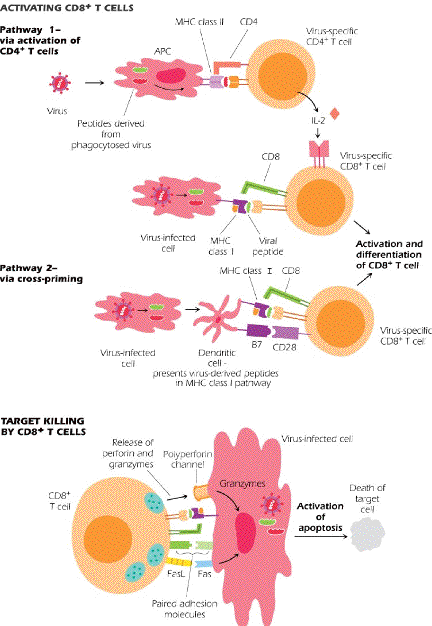
Note that Cytotoxic T Lymphocytes (CTL’s, CD8(+) T-cells) can be activated by two pathways. In both pathways the CTL must recognized antigen which has been processed and presented on MHC class I molecules.
· In the first pathway, CD4(+) T-cells (TH1 cells) provide help in launching the CTL on its way
· In the second pathway, dendritic cells provide the help. The figure indicates that the help is different – this probably prevents the dendritic cell from being killed by the CTL!!??
After activation, the CTL is now capable of killing target cells – that is, cells that are expressing the activating antigen in the context of MHC class I.
There are two killing mechanisms:
· The secretion of enzymes onto the target cell which induce permeability changes and apoptosis – perforin and granzymes
· The interaction of Fas on the CTL with FasL on the target cell activates caspases within the target cell and results in apoptosis.
T-Independent antigens
There are a number of antigens – usually polysaccharide or lipopolysaccharide – with multiple repetitive epitopes, that are capable of activating reactive B-cells without T-cell help.
These antigens generate the production of IgM and there is no apparent immunological memory.